|
中国社会科学院研究生院 摘 要:田螺山遗址作为河姆渡文化的一处重要遗址, 自发掘以来, 其稻作农业的发展程度一直是研究热点, 伴随其发掘和研究的深入, 研究者得出了此阶段生业模式是基于采集渔猎, 稻作农业仍为补充、且水稻尚在驯化过程中的观点。菱角的食用在河姆渡早期阶段出现了一个大幅度的减少, 到河姆渡文化二期晚期阶段几乎不再在人类的日常饮食中占据重要地位, 推测先民们虽同时进行野生资源的采集和水稻种植, 但是其比重已经发生了变化, 更多的开始依赖于早期农业。 关键词:田螺山遗址; 菱角; 生业模式 作为长江下游地区一支重要的新石器晚期考古学文化———河姆渡文化, 自确立以来, 已发现四十余处文化遗址[1]。自河姆渡遗址发掘伊始, 学者们就关注到对其文化生业模式的研究, 究竟是以稻作农业为主还是基于采集渔猎、稻作农业仅为补充?研究者们提出了不同的见解[2]。2001年, 田螺山遗址的发现, 以及其后连续四年的考古发掘, 取得了丰硕的成果, 特别是菱角一类水生可食植物果实的大量遗存, 为河姆渡文化早期生业模式的深入研究提供了丰富的资料。 本文即基于田螺山遗址考古发现的菱角遗存, 通过计量统计和分析, 发现在河姆渡文化早期阶段曾出现过明显的差异, 提出河姆渡文化早期的生业模式可能发生过重大的变化, 不当之处请专家指正。 一、遗址概况 田螺山遗址位于浙江省余姚市三七市镇相村, 西距余姚市区24公里, 地处宁绍平原东部的姚江流域。2004年以来, 浙江省文物考古研究所连续进行了四年的发掘, 累计发掘面积约1000平方米, 发现了十分丰富的河姆渡文化各阶段的遗迹、遗物。由于其特殊的埋藏环境, 使许多有机质遗物得以保存下来, 诸如菱角、栎果、芡实、葫芦、酸枣、稻米等植物遗存以及独木舟、木浆、木质碟形器和干栏式建筑等[3], 为河姆渡文化生业模式的研究提供了十分丰富的实物资料。 依据考古发掘报告[4], 田螺山遗址地层堆积共分为8层。其中第 (3) ~ (8) 层为河姆渡文化地层堆积, 大体对应河姆渡遗址的 (2) ~ (4) 层。发掘者根据土质、土色、叠压次序及出土器物的形态差异, 将遗址分为三个时期的遗存, 分别与河姆渡文化的一、二、三期相对应。其中第 (7) 、 (8) 层为河姆渡文化一期, 上限在距今7000年左右, 下限不早于距今6700~6800年;第 (5) 、 (6) 层为河姆渡文化二期, 年代上限约距今6700~6600年, 第三期则持续到距今6100~6000年[5]。 田螺山出土的植物遗存极为丰富, 根据系统进行过的植物考古学工作[6], 目前所发现的植物种属囊括了六十余科, 多达上百个种属。应田螺山遗址考古发掘领队孙国平先生应允, 笔者有幸参与了田螺山遗址的部分植物考古工作, 主要是对田螺山遗址出土的菱角进行植物考古统计和分析。 表一各层位出土菱角体积总量数据表 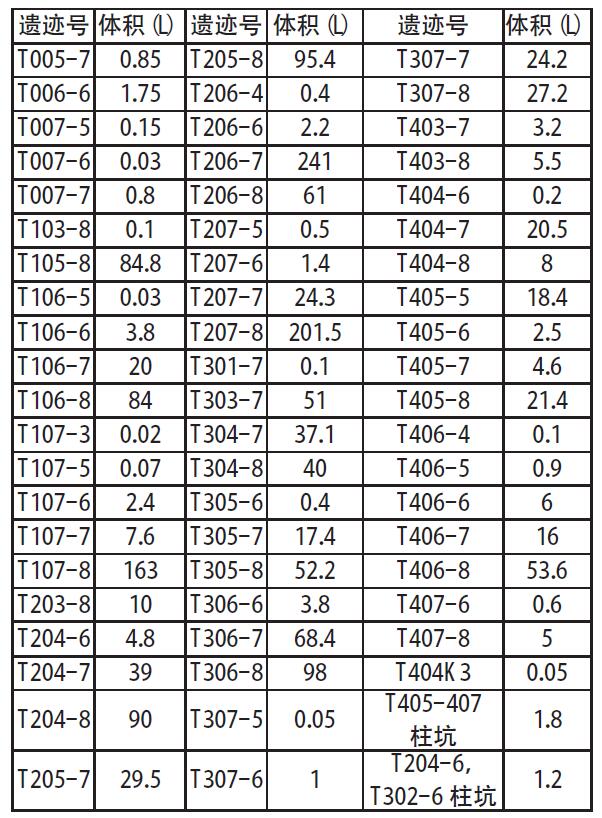 二、采样方法和统计结果 如上所述, 此次工作系将以往尚未进行植物考古工作的菱角样品进行统计, 其中绝大部分属于地层堆积的样品。田螺山遗址的发掘工作十分细致, 浮选土样采集量很大, 这次计量统计工作中共采集了63份样品, 其中2份来自于柱坑, 1份来自于K3, 其余的60份来自于23个探方的不同地层中。这些遗存从年代上来说, 均属于田螺山遗址的第一期遗存和第二期遗存, 即属于河姆渡文化一期和二期的文化遗存。 众所周知, 古代遗址的文化层堆积在同一个遗址中, 并非是均匀分布的, 即便在同一个探方之内, 地层堆积也不完全具有连贯性。同时, 不同层位的堆积由于时间长短的不同和堆积形成过程中的差异, 其文化层厚度也是不相同的。为尽量使统计数据具有科学性, 我们选用的所有63个样品充分考虑了同一个探方地层堆积的连贯性, 主要是统计采集样品的体积和出土菱角的体量。 田螺山遗址所处地下水位较高, 其各类植物遗存大多处于饱水状态, 这也是保存状况较好的主要原因。菱角类遗存虽然出土量较大, 但大多为果皮碎片, 仅有少量的完整个体。因此, 如单纯以菱角个体作为统计单位, 尽管相对简单易行, 但肯定会出现统计结果与实际数量的误差。所以本次统计采取了对每个遗迹单位所有菱角 (包括完整个体和碎片) 的体积进行计量的方法, 这种统计方法尽管也会存在一定的误差, 但就考古发现资料保存现状, 我们认为这种统计方法相对是科学的。在实际操作中, 笔者将每个地层或遗迹单位中的菱角遗存全部收集, 分别利用量筒、烧杯等有刻度的容器进行体积称量, 进行记录, 以便计量, 63份样品的统计结果见表一。 三、统计结果的分析 由表一可以看出, 田螺山遗址的采集样品中, 有的在同一个探方之内地层堆积具有连续性, 如T106即分别采集了 (6) ~ (8) 层的样品, 而有的探方如T005则仅采集了 (7) 层的土样。考虑到地层堆积的连贯性和样品时间的连续性, 因此在讨论时我们选取了相邻的六个探方, 且都具有四个地层连续堆积的样品进行比较 (采样探方关系见图一) 。 各探方不同地层出土菱角数量见表二。 从表二可看出, 不同探方中菱角的统计数据差距较大, 但不同地层之间变化的总趋势却非常一致。遗迹单位中菱角的体积数值在整个田螺山遗址的 (5) ~ (8) 层之间有显著的下降趋势, 各探方中均是在第 (8) 层为最大值, 第 (5) 层为最小值, 下降幅度最大的从201.5L下降至0.5L。在整个下降的过程中, 在某个阶段会发生骤降。在部分探方中这个过程发生在 (6) 、 (7) 层之间, 在部分探方中会发生在5、6层之间。 根据对田螺山遗址的分期, 可将 (7) 、 (8) 层放在一起, 为一期遗存, 对应河姆渡一期文化; (5) 、 (6) 层放在一起, 为二期遗存, 对应河姆渡二期文化。通过一期和二期遗存的比较, 可发现, 田螺山遗址中菱角体积的最大变化发生在河姆渡文化一、二期之间, 几乎呈现出数十倍甚至上百倍的差距 (图二) 。 菱角作为一年生水生草本植物, 其果实富含淀粉质, 每年7~11月均可采收, 是先民们采集经济中食物的重要组成部分。但这种果实并不适宜长期储存, 采摘后的果肉会很快失水萎缩, 甚至丧失其食用性。因此, 这类植物果实并不适宜作为长期的储存类食物。田螺山遗址的菱角样品均采自于地层, 且绝大部分为果壳碎片, 完整菱角和果肉均很少, 除不易保存的原因外, 更大的可能是这些遗存应来自于日常食用的残留, 从而反应出当时菱角在人类食谱中的比重。也就是说在河姆渡一期和二期之间, 人类对菱角的食用出现了大幅度减少的重大变化, 河姆渡一期文化中, 菱角的采集量抑或是食用量居高不下, 而到河姆渡文化二期, 数量锐减。在属于二期的两个层位中, 即从 (6) 层到 (5) 层, 菱角这类食物几乎不见或很少, 即在当时人们的食物中已不再占有重要位置。 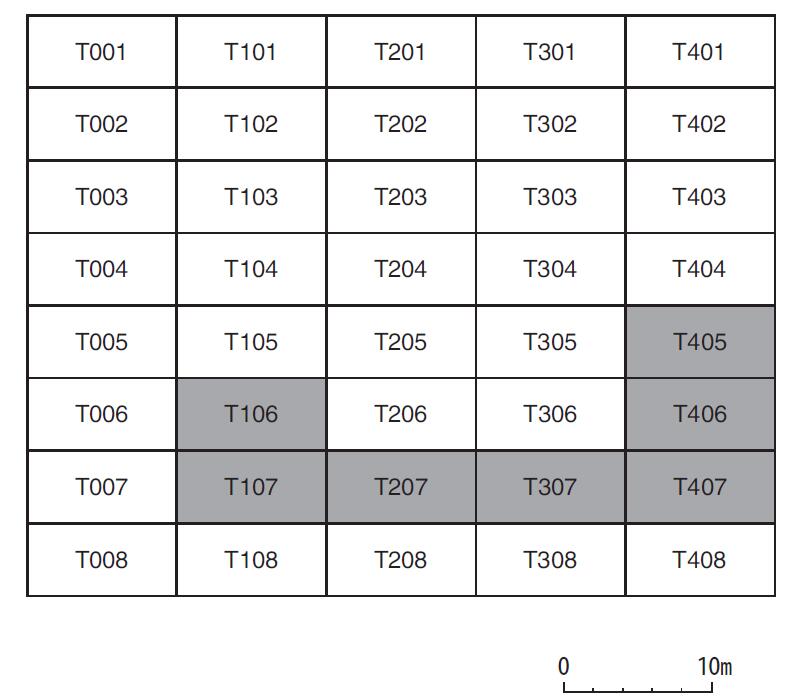 图一采样探方分布图 表二地层出土菱角体积对比表 (单位:L) 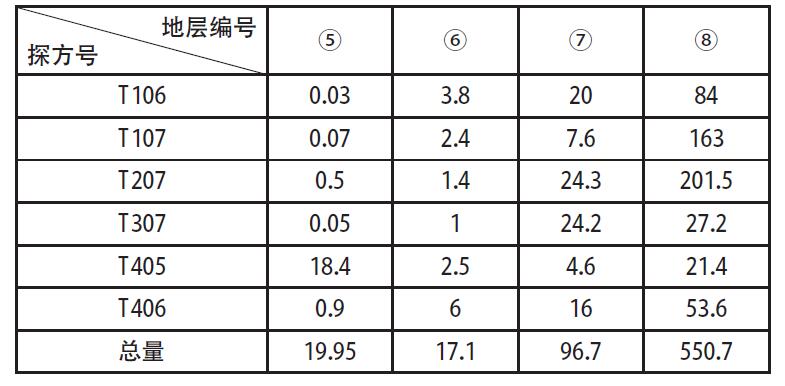 图二 田螺山一、二期菱角体积变化 (单位:L) 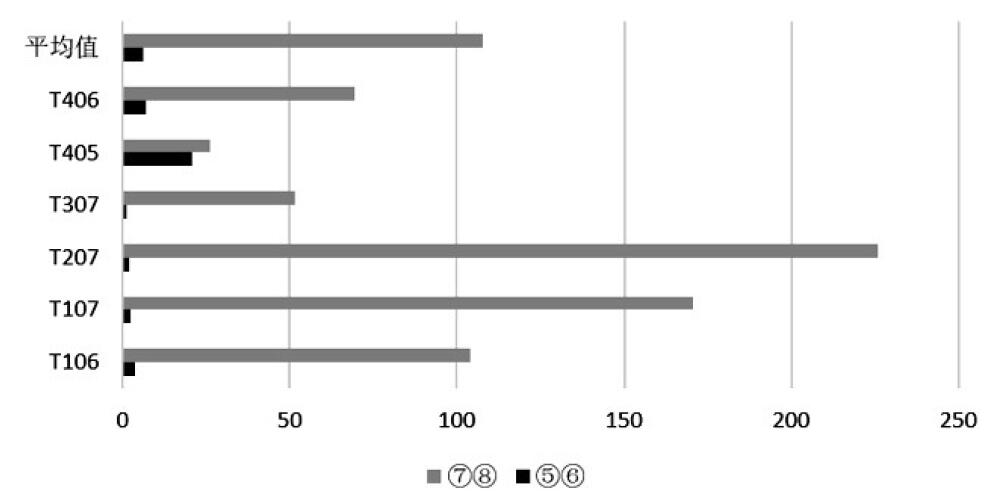 四、相关问题的讨论 从统计结果来看, 田螺山遗址考古发现在河姆渡文化一、二期之间菱角数量存在着锐减。那么, 造成这种现象的原因究竟是什么?河姆渡文化一期大量菱角果壳碎片的存在, 当然与其保存环境比如不同地层堆积水土的酸碱度、地下水位的变化、含水量等有一定关系[7], 但从共存的其他有机类遗物来看, 共存的芡实等食物资源同样存在着下降的趋势, 而某些共存的其他植物遗存如葫芦科、莎草科等, 却无论在绝对数量还是出土概率上却都并没有下降趋势, 甚至个别 (莎草科、壳斗科) 呈现出上升趋势[8]。这种情况反应, 保存环境虽然会对菱角类遗物的保存现状产生一定影响, 但不会是决定性作用。因此, 大量菱角果壳碎片的存在主要是人为的原因, 也即为, 菱角在河姆渡一期文化时期, 较之于二期文化更广泛地作为当时人们的食物使用, 果壳碎片的存在当是人们食用之后的残留物。不同层位菱角数量上的巨变, 可能反应了人们食物取向出现了变化。 曾有学者推测, 造成菱角这种锐减的现象也有可能与淡水资源的减少或者是菱角的过度开采导致数量上的大量减少有关[9]。诚然, 根据对孢粉、沉积物和地球化学的研究结果[10], 河姆渡遗址群确是根植于海退而发展起来, 在河姆渡文化之前、河姆渡文化中期及河姆渡文化之后, 均出现海平面较高的时期, 对文化的进程造成了比较大的影响。两次大规模的沼泽化, 使得淡水区域有所扩张和紧缩。但就本文讨论依据的田螺山遗址 (5) ~ (8) 层, 也就是河姆渡文化一、二期之间, 田螺山遗址几乎没有受到海水的影响, 海水降低到海拔两米以下[11], 气候是比较稳定的, 大致相当于现今福州一带, 明显比今天湿润温暖。在距今6000年后气候的暖湿程度才逐步降低, 此时期已属于村落化定居和人口较为稠密的时期。 来自动物考古的研究也可支持这一观点, 从第 (8) 层到第 (3) 层淡水鱼类骨骼遗存的数量呈现上升趋势[12], 与同为淡水植食资源的菱角和芡实的数量改变正好相反, 在田螺山一期和二期之间也保持着较为稳定的数量。这说明, 在此时期内, 人类并没有放弃对淡水中的动物资源的开发, 仅仅是减少了对淡水植物资源的采集和利用。 由此可见, 环境对于菱角数量大幅度减少的影响是非常有限的, 更多的原因应归于生业模式的改变。 与菱角同为淡水植物资源的, 还有睡莲科芡属的芡实。芡实也为一年生的水生植物, 常与莲、菱混生。根据已发表的材料[13], 芡实的数量在田螺山的一期和二期遗存之间也呈现出了相同的趋势, 从一期到二期发生了大幅度的减少, 到第三期几乎不占比重。其出土概率也在从一期的几乎100%降至二期的60%。 与此同时, 在田螺山遗址中, 人们对壳斗科植物即栎果的利用并没有大幅度增加。栎果即所谓的“橡子”, 富含淀粉, 且有一定的脂肪含量, 可作为长期储存的食用资源。栎果在田螺山遗址一期、二期的利用保持着较为稳定的状态, 到三期后, 对于栎果的利用也开始减少[14]。这说明人们在放弃了对菱的大量采集利用之后, 也并没有从此转向对林缘植物———栎果大量消费上。由此可见, 对于菱角的采集利用的减少, 也并非是源于对其他野生资源的大量开发。 由此, 从田螺山遗址考古出土菱角数量的巨大变化, 我们有理由推测, 河姆渡文化时期先民的生业模式出现了变化, 可能是逐渐减少了对野生植物资源的摄入, 从而转到了对农业的开发上。 自上世纪河姆渡遗址发现以来, 河姆渡文化稻作农业一直是研究热点。随着考古新资料的不断发现和研究的不断深入, 基本已修正了河姆渡文化发现初期认为河姆渡文化是以稻作农业为主体的观点, 有学者提出了在河姆渡文化时期, 稻作农业仍处于较为初期的发展阶段, 水稻正处于驯化的过程中, 河姆渡仍是一个“以坚果采集和渔猎为主要生计模式的早期社会形态”[15]。虽然从数量上来说, 发现水稻的数量并不是持续上升, 而是略有波动, 但是通过植物考古的方法, 发现水稻驯化的特征却不断加强[16]。如果驯化被定义为“在人类行为干预下, 在遗传学和形态学意义上植物发生的变化”[17], 那么起码说明这段持续的时间内, 人类的水稻的种植行为和人为干预是一直在加强的。 淀粉是人类营养来源中不可或缺的一部分, 就田螺山先民的食谱来看, 菱角、芡实、栎果、水稻均为其主要的淀粉类食物来源。通过以往对这几种植物遗存的研究, 我们可看出, 随着对菱角的利用的下降, 可能是由于对水稻利用的加强。根据对田螺山菱角的定量统计, 可以推测, 到河姆渡文化二期, 菱角在人类食谱中已经不占据重要地位, 但是也并没有转向大量的对林缘植食资源的开发利用。所以有理由这样认为, 田螺山的先民们在河姆渡文化时期, 虽然同时进行野生资源的采集和水稻种植, 但是其比重已经发生了变化, 更多的开始依赖于早期农业。 注释: [1]冯小妮、高蒙河:《宁绍地区早期遗址群的量化分析》, 《东南文化》2004第6期;王海明:《河姆渡遗址与河姆渡文化》, 《东南文化》2000第7期。 [2]浙江省文考古研究所:《河姆渡--新石器时代遗址考古发掘报告》, 文物出版社, 2003年。 [3]浙江省文物考古研究所:《田螺山遗址第一阶段 (2004~2008年) 考古工作概述》, 见:北京大学考古文博学院, 浙江省文物考古研究所:《田螺山遗址自然遗存综合研究》, 文物出版社, 2011年。 [4]同[3]。 [5]潘艳:《长江三角洲于钱塘江流域距今10000~6000的资源生产:植物考古与人类生态学研究》, 复旦大学博士学位论文, 2011年。王淑云、莫多闻、孙国平、史辰羲、李明霖、郑云飞、毛龙江:《浙江余姚田螺山遗址古人类活动的环境背景分析---植硅体、硅藻等化石证》, 《第四纪研究》, 2010年第2期。 [6]傅稻镰、秦岭、赵志军、郑云飞、细谷葵、陈旭高、孙国平:《田螺山遗址的植物考古学分析:野生植物资源采集、水稻栽培和水稻驯化的形态学观察》, 见:北京大学考古文博学院, 浙江省文物考古研究所:《田螺山遗址自然遗存综合研究》, 文物出版社, 2011年。潘艳:《长江三角洲于钱塘江流域距今10000~6000的资源生产:植物考古与人类生态学研究》, 复旦大学博士学位论文, 2011年。 [7]郭怡、李泊提、孙国平、奚三彩:《田螺山遗址古菱角埋藏环境调查与保护对策的初步研究》, 《南方文物》2013年第1期。 [8]同[5]。 [9]郑云飞、陈旭高、孙国平:《田螺山遗址出土植物种子反映的食物生产活动》, 见:北京大学考古文博学院, 浙江省文物考古研究所:《田螺山遗址自然遗存综合研究》, 文物出版社, 2011年。 [10]莫多闻、孙国平、史辰羲、李明霖、王淑云、郑云飞、毛龙江:《浙江田螺山遗址及河姆渡文化环境背景探讨》, 见:北京大学考古文博学院, 浙江省文物考古研究所:《田螺山遗址自然遗存综合研究》, 文物出版社, 2011年。温证聪、孙国平、谢柳娟、孙永革:《河姆渡文化田螺山遗址古土壤有机质的地球化学特征及其意义》, 《地球化学》, 2014第2期。 [11]王淑云、莫多闻、孙国平、史辰羲、李明霖、郑云飞、毛龙江:《浙江余姚田螺山遗址古人类活动的环境背景分析--植硅体、硅藻等化石证》, 《第四纪研究》, 2010年第2期。金原正明、郑云飞:《田螺山遗址的硅藻、花粉和寄生虫卵分析》, 见:北京大学考古文博学院, 浙江省文物考古研究所:《田螺山遗址自然遗存综合研究》, 文物出版社, 2011年。 [12]同[9]。 [13]同[6]。 [14]潘艳:《长江三角洲于钱塘江流域距今10000~6000的资源生产:植物考古与人类生态学研究》, 复旦大学博士学位论文, 2011年。 [15]傅稻镰、秦岭、赵志军、郑云飞、细谷葵、陈旭高、孙国平:《田螺山遗址的植物考古学分析:野生植物资源采集、水稻栽培和水稻驯化的形态学观察》, 见:北京大学考古文博学院, 浙江省文物考古研究所:《田螺山遗址自然遗存综合研究》, 文物出版社, 2011年。 [16]同[6]。 [17]傅稻镰、秦岭、胡雅琴 (译) :《稻作农业起源研究中的植物考古学》;《南方文物》, 2009年第3期。 (责任编辑:admin) |