|
大豆是世界重要经济作物和粮食作物之一,因其高蛋白、高脂肪含量成为粮油和饲料来源。在植物分类学中,大豆属包括两个种,即野生大豆和栽培大豆。野生大豆为栽培大豆的近缘野生祖先,主要分布于中国、朝鲜半岛、日本列岛和俄罗斯远东地区。细胞生物学和分子生物学证据均支持野生大豆经自然及人工选择逐步进化为栽培大豆。 野生大豆和栽培大豆基因序列的对比分析表明,大豆的栽培历史十分复杂。黄河中下游地区长期以来被视为大豆起源地之一。这里迄今年代最早的大豆属遗存,见于新石器时代早期(距今9000—7500年)。至迟在龙山时代(距今4500—4000年),已经开始从尺寸上分化为大和小两组,暗示着野生大豆和栽培大豆的共存。 从考古资料看,与新石器时代早期的炭化大豆属遗存伴出的,还有粟、黍、水稻等栽培作物,它们与陶器、房屋、灰坑等遗存共同指向一种相对稳定的聚落生活。此时的炭化大豆遗存,出土频率较高,推测古人已经开始介入野生大豆的生境,并导致早期野生大豆发生某些特定的性状变化,只是没有体现在尺寸这一特征上。显然,大豆尺寸的变化在人工栽培初期十分模糊,种粒变大这一性状的改变,是驯化较晚阶段才发生的。而其他栽培相关特征,如豆荚开裂程度、籽粒硬实性等,在缺少古代DNA数据支持的前提下,则难以从考古材料中辨识出来。 遗传学研究利用QTL和GWAS等技术可以探讨大豆脂肪与蛋白质含量的变化。就种子本身成分而言,进化程度低、野生性状典型的野生大豆蛋白质含量高,大豆的蛋白质含量与百粒重、脂肪含量呈负相关。伴随X-CT高精度无损扫描技术的引入,我们可以对炭化大豆种子的内部结构进行细致的观察和测量。炭化大豆内部有着大小不一的孔洞结构,这些孔洞能否与种子的成分之间建立关联,从而成为大豆起源研究新的突破点? 沿着这一思路,山东大学研究团队使用同步辐射X射线显微成像技术,对现代和古代炭化大豆孔洞结构作了测量和三维重建。实验结果表明,炭化大豆的孔洞结构具有独特性,且与油滴的关系非常密切。油滴沿子叶流动并聚合为大油滴,一经炭化,大油滴消失,最终形成炭化种子内部的孔洞结构。对已测定蛋白质和脂肪含量的三组现代样品模拟炭化后采集数据显示,蛋白质与油脂的总体含量和炭化大豆孔洞的各组数量比之间存在线性对应关系。高油脂可能促使更多较小孔洞的生成;反之,高蛋白导致了较大孔洞的产生。 将这一关联运用到黄河中下游地区四处遗址出土的七份考古样品中(如图),每粒种子的孔洞分布情况,体现在孔洞体积比(纵轴)及其数量排序(横轴)上。孔洞体积比,是指孔洞体积除以种子体积;数量排序,即每粒大豆的孔洞由大及小逐个排序。每条曲线代表一粒大豆,曲线上的点代表孔洞。 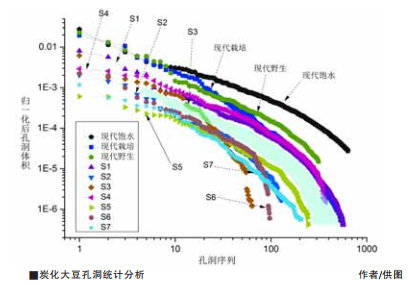 如图所示,随着时间的推进,大豆含油量趋于增加。三粒现代样品和考古样品S1(唐代,西金城)和S4(龙山,莘冢集),称其为组1,其孔洞体积分布明显高于其他样品。而S2—S3(商代,大辛庄)、S5—S6(龙山文化,西金城)和S7(后李文化,月庄),称其为组2,其孔洞体积分布则靠下。浅色空白区域将组1和组2明显划分为两个时代,两者之间的时代差距达2000年以上(S4除外),含油量涨幅更大(右侧曲线更为发散),显示含油量提升明显。 大辛庄遗址商代炭化大豆尺寸明显可分为小和大两组,或指向野生种和栽培种的分化。我们将大辛庄遗址出土的一大一小两粒样品S2和S3进行比较,以观察大豆内部成分差异与尺寸之间是否存在对应关系。S2和S3尺寸大小差别明显,推测S2系栽培种,S3为野生种。从CT扫描结果看,S3比S2含油量低,而蛋白质含量高于S2。S2含油量高于S3,蛋白质含量比S3低。这一结果支持了S2系栽培种、S3为野生种的判断。成分分析与尺寸判断结果一致。 样品S5和S6是来自西金城遗址的龙山时代遗存。二者个体都比较大,甚至都大于大辛庄的商代样品。根据李炅娥等人的统计,炭化大豆长度和宽度数值具有较强的线性相关性。因此,长度数值是区分大小的关键数值。据此可视,S5个体大于S6。尽管孔洞序数(横轴)在20—60范围内区分不明显,但在序数80以后,S5样品中具有更多的小孔洞。这暗示了S5要比S6含油量高。成分分析与尺寸判断结果相符。此外,年代最早的后李文化样品S7,其含油量甚至高于现代野生大豆。 这表明,黄河中下游地区先民选择培育大豆的历史,最迟距今7500年前就已经发生,且更倾向于选择脂肪含量高的个体。这一发现,弥补了之前尺寸分析所不能观察到的人类对野生大豆的早期干预进程,也部分回答了李炅娥等根据尺寸分析揭示的日本列岛大粒大豆出现时间(距今5000年)早于中国这一问题。该研究显示出同步辐射X-CT手段在探索作物驯化方面的巨大潜力。 (作者单位:山东大学文化遗产研究院) (责任编辑:admin) |